背景介绍
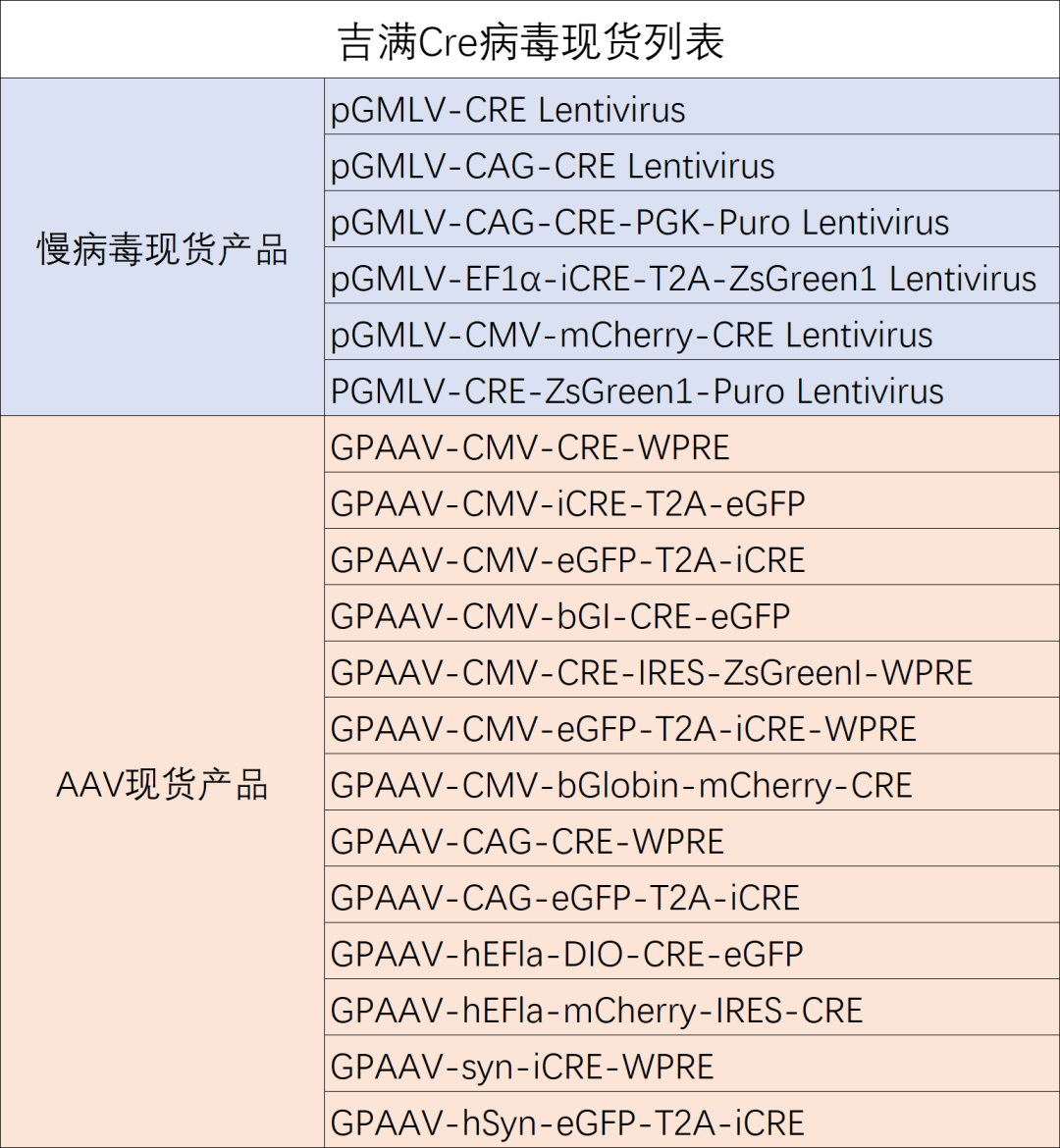
Cre 重组酶(Cyclization Recombination Enzyme, 即环化重组酶),是细菌噬菌体P1的I型拓扑异构酶。其基因编码区序列全长1029bp,为38kDa大小的、由343个氨基酸组成的多肽单体蛋白。Cre重组酶的C-末端结构域包含催化活性位点,能够催化DNA分子中特定位点之间的重组。而且与限制酶相似,能识别特异的DNA序列,即loxP位点,使loxP位点间的基因序列被删除或重组。
LoxP序列 locus of X (cross)-over in P1),是位于P1噬菌体中长度为34bp的一段序列,由两个13bp的反向回文序列和8bp的中间间隔序列共同组成,反向回文序列是Cre重组酶的识别和结合区域,间隔序列决定LoxP序列的方向。loxP有多种突变类型,多种突变类型之间不交叉识别。

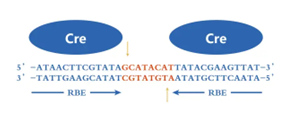
Cre-loxP系统基因重组原理
一般而言,当细胞基因组内存在两个LoxP位点时,Cre重组酶会诱导两个LoxP位点间的序列发生重组。
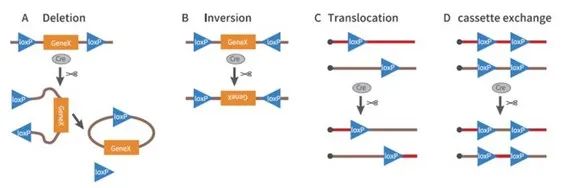
Cre-loxP诱导基因重组的方式
Cre重组酶介导两个LoxP位点间的重组是一个动态、可逆的过程,重组的结果取决于两个loxP位点的方向,主要有以下情况:
1、删除(Deletion)
如果两个LoxP位点位于一条DNA链上,且方向相同,Cre重组酶能有效切除两个LoxP位点间的序列
2、倒转(Inversion)
如果两个LoxP位点位于一条DNA链上,但方向相反,Cre重组酶能导致两个LoxP位点间的序列倒转
3、易位(Translocation)
如果两个LoxP位点分别位于两条不同的DNA链或染色体上,Cre酶能介导两条DNA链的交换或染色体易位,即基因转座
4、序列互换(Cassette exchange)
如果四个loxP位点分别位于两条不同的DNA链或染色体上,Cre重组酶能诱导loxP间的序列互换(cassette exchange)
两种策略实现Cre依赖的基因表达
1、DIO序列策略
DIO序列策略的关键在于引入两对不同的lox位点:LoxP和Lox2272。在Cre酶的作用下,不论是识别LoxP还是Lox2272位点,第一步都可以实现目标基因的反转。反转会使两个loxP位点或者Lox2272位点变成同向。第二步在Cre酶的作用下,剪切删除一个LoxP位点和一个Lox2272位点,进而实现基因的稳定表达。
2、LSL序列策略
LSL序列策略是将LoxP和转录中止盒插入启动子和目的基因中间,构成转录“终止盒”(Lox-stop-Lox),这个“终止盒”能够终止基因的转录。在没有Cre酶的作用下,转录终止盒下游靶基因完全不表达。但是如果细胞中有Cre酶,终止盒会被Cre酶切除,从而实现组织特异性的基因过表达。
Cre/loxP条件性重组改造
经过现代基因工程方法对Cre和LoxP元件的改造,Cre-LoxP系统实现了更加丰富的条件性重组策略。
1、对Cre元件的改造
提高Cre重组酶活性;构建可诱导的方式
2、对loxP元件的改造
对间隔区和回文序列进行突变,突变后的序列依然能被Cre重组酶识别和重组,但是突变的loxP序列必须和同样突变的loxP序列匹配介导基因重组,在同一Cre重组酶的作用下,可以实现多序列的基因重组,产生非常多元的重组结果。下表为常见的loxP类型:

LoxP位点及其突变体位点
Cre-loxP系统的优点
1、高效性
Cre重组酶与具有loxP位点的DNA片段形成复合物之后,可以提供足够的能力引发之后的DNA重组过程,重组过程简约高效;重组不受切除片段长短及位置影响,可以在活体动物体内实现,可随细胞分裂和动物传代稳定遗传。
2、特异性强
loxP位点是一段含回文序列结构和中间有间隔的34bp元件,这种结构保证了loxP序列的唯一性,从而保证基因重组的特异性很强;在动物模型中至今未发现Loxp位点之外的非特异性重组,只在体外实验中有少量报道。
3、应用范围广
Cre重组酶是一种比较稳定的蛋白质,可以在生物体不同的组织、不同的生理条件下发挥作用,所以说Cre-loxP系统的可应用范围非常广。
4、快速性
动物实验已证实在受精卵分裂前的短时间内,Cre介导的特异性重组便会完成。
5、时空特异性
如果将Cre结构基因置于某一特定启动子或其他调节基因控制之下,便会实现Cre酶蛋白在特定组织和特定时间的可控型表达,继而限定了重组发生的时空性。

Cre-LoxP重组系统在转基因中的应用
病毒依赖的基因重组的优势
1、更强的区域特异性
由于病毒可以通过局部注射的方式保证区域特异性感染,再加上驱动Cre基因的特异性启动子,能够实现更强的区域和细胞特异性的基因重组。
2、更少的花费
购买转基因动物的费用一般是比较昂贵的,而且转基因动物的饲养、基因型鉴定都需要不少的人力、物力,而病毒的制备、保存和注射所花的费用相对来说是比较少的。
3、更短的实验周期